


 11 марта, 2014
11 марта, 2014  Андрей Сорокин
Андрей Сорокин 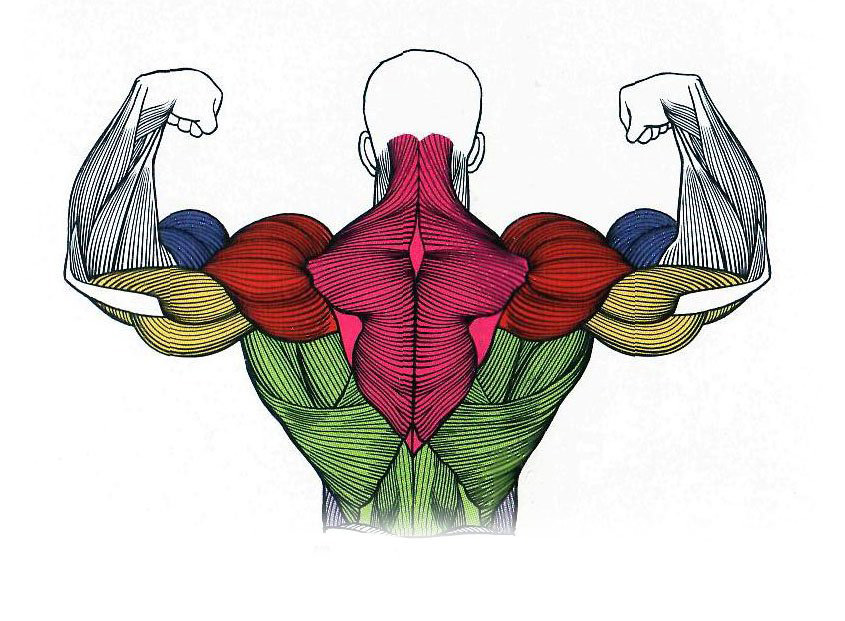 Энергетика мышечной деятельности.
Энергетика мышечной деятельности.
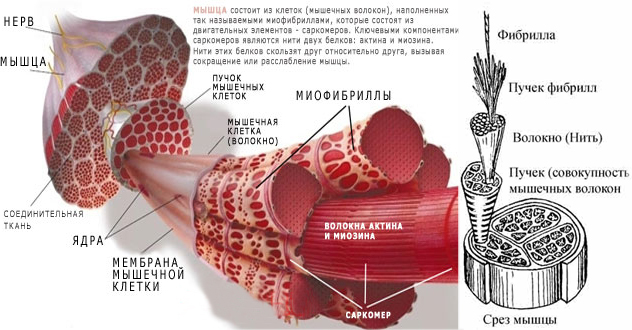
Одно мышечное волокно может содержать 15 миллиардов толстых нитей. При том, что мышечные волокна активно сокращаются, в каждой толстой нити распадаются примерно 2500 молекул АТФ (нуклеотид, играющий важную роль в обмене энергии и веществ в организме) в секунду. Даже небольшие скелетные мышцы содержат тысячи мышечных волокон.
Основной функцией АТФ является передача энергии из одного места в другое, а не долгосрочное хранение энергии. В состоянии покоя скелетные мышечные волокна производят больше АТФ, чем они нуждается. В этих условиях АТФ передает энергию креатина. Креатин представляет собой небольшую молекулу, которую мышечные клетки собирают из фрагментов аминокислот. Передача энергии создает еще одно высокоэнергетическое соединение креатин фосфата (КФ).
АТФ + креатин АДФ + креатин фосфат
Во время мышечного сокращения происходит разрыв соединений АТФ, в результате этого образуется аденозиндифосфат (АДФ). Энергия, запасенная в креатинфосфате затем используется для «перезарядки» АДФ, превращая его обратно в АТФ через обратную реакцию.
 АДФ + креатин фосфат + креатин
АДФ + креатин фосфат + креатин
Фермент креатинфосфокиназа (КФК) облегчает эту реакцию. Когда мышечные клетки повреждены, происходит утечка КФК через клеточные мембраны в кровоток. Таким образом, высокая концентрация в крови КФК обычно указывает на серьезное повреждение мышц. Отдыхающие скелетные мышечные волокна содержат примерно в шесть раз больше креатин фосфата как АТФ. Но когда мышечные волокна переживают устойчивое напряжение, эти энергетические запасы будут исчерпаны всего лишь примерно за 15 секунд. Мышечные волокна должны затем полагаться на другие механизмы для преобразования АДФ в АТФ.
Большинство клеток в организме генерируют АТФ через аэробный метаболизм в митохондриях и через гликолиз в цитоплазме. Аэробный метаболизм (сопровождающийся потреблением кислорода) обычно обеспечивает 95 % АТФ в покоящейся клетке. В этом процессе митохондрии поглощают кислород, АДФ, ионы фосфата и органические субстраты от окружающей цитоплазмы. Субстраты затем вводят цикл трикарбоновых кислот (также известный как цикл лимонной кислоты или цикл Кребса), ферментативный путь, который разрушает органические 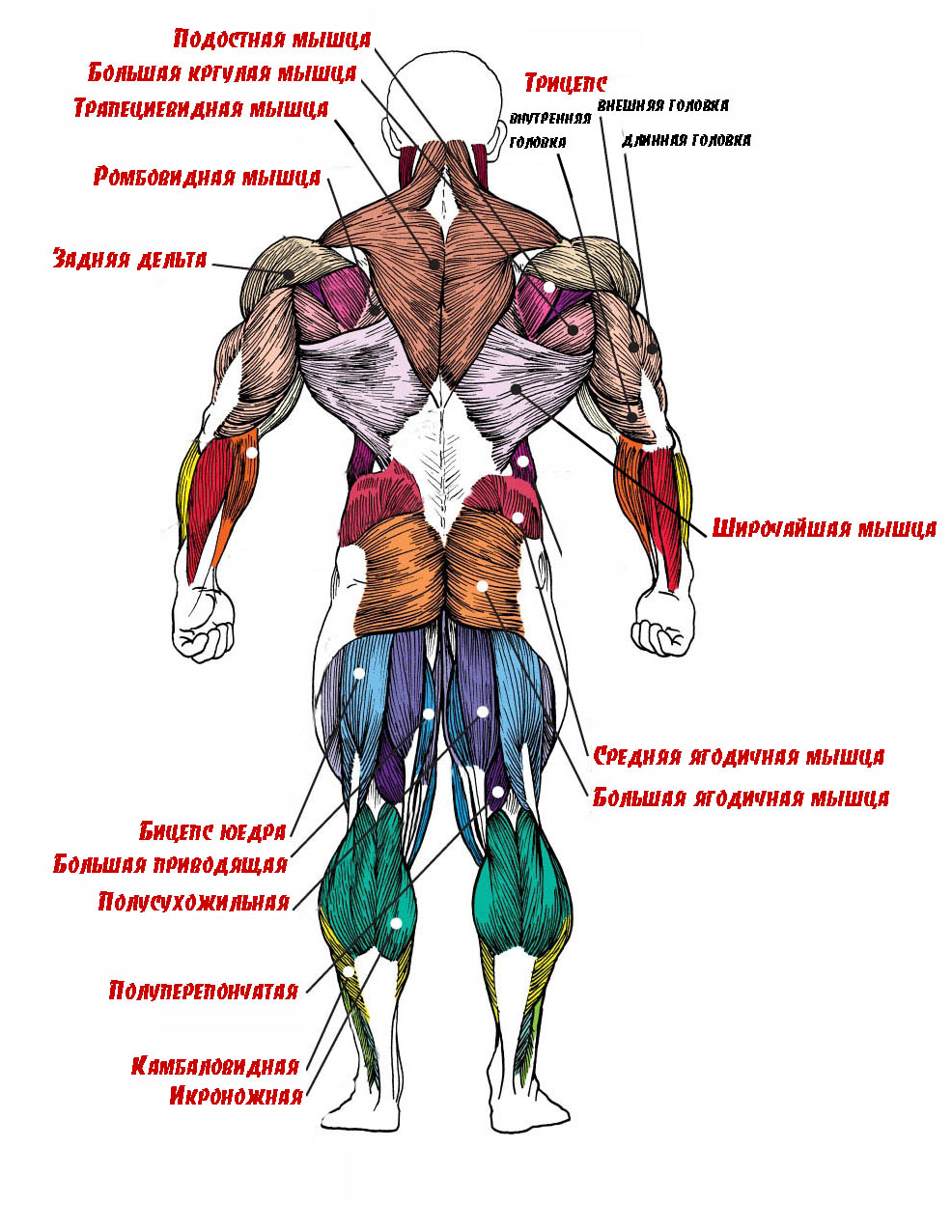 молекулы. Атомы углерода выпускаются, как двуокись углерода, атомы водорода курсируются дыхательными ферментами во внутренней митохондриальной мембране, где их электроны удаляются. После серии промежуточных шагов, протоны и электроны сочетаются с кислородом и образованием воды. В этом эффективном процессе высвобождается большое количество энергии и используется для создания АТФ.
молекулы. Атомы углерода выпускаются, как двуокись углерода, атомы водорода курсируются дыхательными ферментами во внутренней митохондриальной мембране, где их электроны удаляются. После серии промежуточных шагов, протоны и электроны сочетаются с кислородом и образованием воды. В этом эффективном процессе высвобождается большое количество энергии и используется для создания АТФ.
Отдыхающие скелетные мышечные волокна полагаются почти исключительно на аэробный метаболизм жирных кислот, чтобы генерировать АТФ. Когда мышца начинает сжиматься, митохондрии начинают разрушение молекулы пировиноградной кислоты вместо жирных кислот. Пировиноградная кислота обеспечивается ферментативным путем гликолиза. Гликолиз является распадом глюкозы до пировиноградной кислоты в цитоплазме клетки. Этот процесс называется анаэробным, потому что он не требует кислорода. Гликолиз обеспечивает увеличение АТФ и генерирует 2 молекулы пировиноградной кислоты из каждой молекулы глюкозы. АТФ образуется в процессе гликолиза. Поскольку гликолиз может протекать в отсутствие кислорода, он может быть особенно важен, когда наличие кислорода ограничивает скорость производства митохондриальной АТФ. В большинстве скелетных мышц гликолиз является основным источником АТФ во время пиковых периодов активности. Расщепление 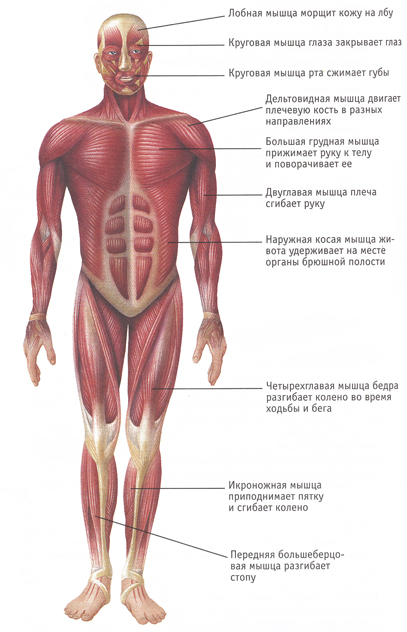 глюкозы в этих условиях происходит в основном из резервов гликогена в саркоплазме. Гликоген представляет собой полисахарид цепочек молекул глюкозы. Типичные волокна скелетных мышц содержат большие запасы гликогена, которые могут составлять 1,5 % от общего веса мышц.
глюкозы в этих условиях происходит в основном из резервов гликогена в саркоплазме. Гликоген представляет собой полисахарид цепочек молекул глюкозы. Типичные волокна скелетных мышц содержат большие запасы гликогена, которые могут составлять 1,5 % от общего веса мышц.
Энергопотребление и уровень мышечной активности.
В скелетных мышцах, при нахождении их в состоянии покоя, спрос на АТФ является низким. Более чем достаточно доступного кислорода митохондриям для удовлетворения этого спроса, в итоге они производят избыток АТФ. Дополнительный АТФ используется для создания запасов гликогена. Отдыхающие мышечные волокна поглощают жирные кислоты и глюкозу, которые доставляются кровотоком. Жирные кислоты расщепляются в митохондриях и АТФ генерируется для преобразования креатина в креатин фосфат и глюкозы в гликоген.
При умеренных уровнях физической активности увеличивается потребность в АТФ. Этот спрос удовлетворяется за счет митохондрий, когда скорость производства митохондриальной АТФ повышается, что увеличивает скорость потребления кислорода. Наличие кислорода не является ограничивающим фактором, потому что кислород может диффундировать (соединяться, смешиваться) в мышечном волокне достаточно быстро, чтобы удовлетворить митохондриальные потребности. Скелетные мышцы в этот момент зависят главным образом от аэробного метаболизма пировиноградной кислоты, чтобы генерировать АТФ. Пировиноградная кислота образуется в процессе гликолиза, который расщепляет молекулы глюкозы, полученные из 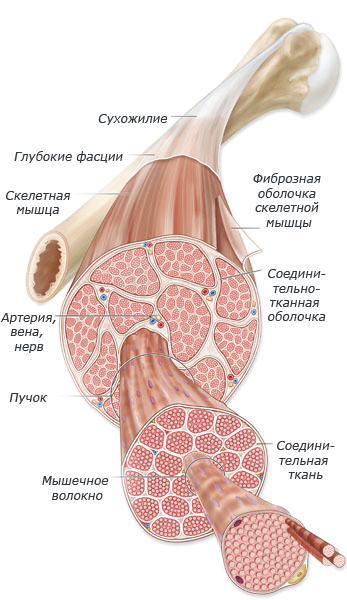 гликогена в мышечных волокнах. Если запасы гликогена низки, мышечное волокно может также расщипить и другие субстраты, такие как липиды или аминокислоты. Пока спрос на АТФ может быть удовлетворен путем митохондриальной активности, обеспечивание АТФ гликолизом остается незначительным в общем энергетическом процессе мышечного волокна.
гликогена в мышечных волокнах. Если запасы гликогена низки, мышечное волокно может также расщипить и другие субстраты, такие как липиды или аминокислоты. Пока спрос на АТФ может быть удовлетворен путем митохондриальной активности, обеспечивание АТФ гликолизом остается незначительным в общем энергетическом процессе мышечного волокна.
В пиковые уровни активности требуется много АТФ, в результате чего производство АТФ в митохондриях возрастает до максимума. Это максимальная скорость определяется наличием кислорода, а кислород не может диффундировать в мышечных волоконах достаточно быстро, чтобы дать возможность митохондрии для образования требуемого АТФ. В пиковых уровнях нагрузки митохондриальная активность может обеспечить лишь около одной трети от необходимого АТФ. Остальная часть приходится на гликолиз.
Когда гликолиз производит пировиноградную кислоту быстрее, чем она может быть использована в митохондрии, увеличивается уровень пировиноградной кислоты в саркоплазме. В этих условиях пировиноградная кислота превращается в молочную кислоту.
Анаэробный процесс гликолиза позволяет клетке генерировать дополнительный АТФ, когда митохондрии не в состоянии удовлетворить текущие потребности в энергии. Тем не менее, производство анаэробной энергии имеет свои недостатки:
Молочная кислота представляет собой органическую кислоту, которая в жидкостях организма  диссоциирует на ионы водорода и отрицательно заряженного иона лактата. Таким образом, производство молочной кислоты может привести к снижению внутриклеточного рН. Буферы в саркоплазме могут противостоять рН сдвигам, но эти защиты ограничены. В конце концов изменения рН будут изменять функциональные характеристики ключевых ферментов.
диссоциирует на ионы водорода и отрицательно заряженного иона лактата. Таким образом, производство молочной кислоты может привести к снижению внутриклеточного рН. Буферы в саркоплазме могут противостоять рН сдвигам, но эти защиты ограничены. В конце концов изменения рН будут изменять функциональные характеристики ключевых ферментов.
Гликолиз является относительно неэффективным способом для генерации АТФ. В анаэробных условиях каждая молекула глюкозы генерирует 2 молекулы пировиноградной кислоты, которые превращаются в молочную кислоту. В свою очередь, клетка получает 2 молекулы АТФ через гликолиз. Если бы те молекулы пировиноградной кислоты катаболизировались бы аэробным способом в митохондриях, клетка получила бы 34 дополнительных молекул АТФ.
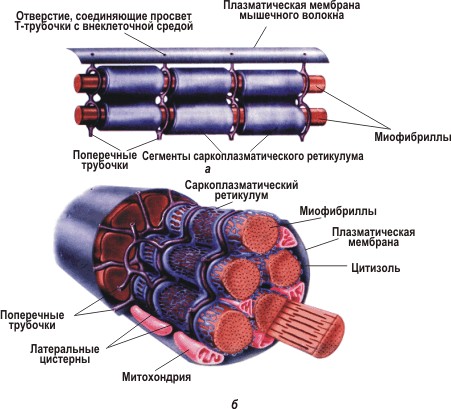
Мышечная усталость. Скелетные мышечные волокна устают тогда, когда они больше не могут сокращаться, несмотря на продолжение нервного импульса. Причина мышечной усталости изменяется в зависимости от уровня активности мышц. После коротких пиковых уровней деятельности, например таких, как 100-метровый забег на время, усталость может быть 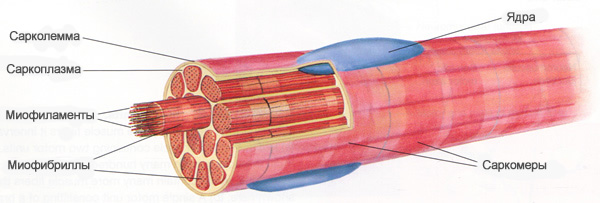 результатом исчерпания запасов АТФ или от падения рН, который сопровождается накоплением молочной кислоты. После длительных напряжений, таких как марафон, усталость может включать физические повреждения саркоплазматического ретикулума, что мешает регуляции внутриклеточных + концентрации ионов Ca2. Мышечная усталость накапливается и последствия этого становятся более выраженными, поскольку все больше мышечных волокон начинают задействоваться этим состоянием. Результатом является постепенное снижение возможностей всех скелетных мышц.
результатом исчерпания запасов АТФ или от падения рН, который сопровождается накоплением молочной кислоты. После длительных напряжений, таких как марафон, усталость может включать физические повреждения саркоплазматического ретикулума, что мешает регуляции внутриклеточных + концентрации ионов Ca2. Мышечная усталость накапливается и последствия этого становятся более выраженными, поскольку все больше мышечных волокон начинают задействоваться этим состоянием. Результатом является постепенное снижение возможностей всех скелетных мышц.
Если мышечное волокно сокращается при умеренных уровнях и запросы АТФ могут быть удовлетворены через аэробный метаболизм, усталость не произойдет, пока запасы гликогена, липидов и аминокислот не будут исчерпаны. Этот тип усталости происходит с мышцами спортсменов долгих нагрузок, таких как марафонцы, после нескольких часов забегов на длинные дистанции.
Когда мышца производит внезапный, интенсивный всплеск активности на пиковых уровнях, большая часть АТФ обеспечивается путем гликолиза. После нескольких секунд до минуты, повышение уровня молочной кислоты снижает рН тканей и мышцы больше не могут функционировать нормально. Спортсмены, которые испытывают быстрые мощные нагрузки, такие как спринтеры в 100-метровом забеге, получают как раз этот тип мышечной усталости.
 Для нормальной функциональности мышц требуется: 1) существенные внутриклеточные энергетические запасы, 2) нормальное кровообращение и 3) нормальная концентрация кислорода в крови. Все, что препятствует одному или более из этих факторов, будет способствовать преждевременной усталости мышц. Например, снижение кровотока от тесной одежды, расстройство кровообращения или потеря крови замедляет доставку кислорода и питательных веществ, ускоряя при этом накопление молочной кислоты, а также способствует мышечной усталости.
Для нормальной функциональности мышц требуется: 1) существенные внутриклеточные энергетические запасы, 2) нормальное кровообращение и 3) нормальная концентрация кислорода в крови. Все, что препятствует одному или более из этих факторов, будет способствовать преждевременной усталости мышц. Например, снижение кровотока от тесной одежды, расстройство кровообращения или потеря крови замедляет доставку кислорода и питательных веществ, ускоряя при этом накопление молочной кислоты, а также способствует мышечной усталости.
Восстановительный период. При сокращении мышечных волокон условия в саркоплазме меняются. Потребляются энергетические запасы, выделяется тепло и, если сокращение было пиковым, генерируется молочная. В период восстановления, условия в мышечных волокнах возвращаются к нормальным. Может занять несколько часов для того,чтобы мышечные волокона оправились от периода умеренной активности. После длительной деятельности на более высоких уровнях активности, полное восстановление может занять неделю. В период восстановления, когда кислорода имеется в изобилии, молочная кислота может быть переработана путем конвертации обратно в пировиноградную кислоту. Пировиноградная кислота может быть использована или митохондрией для генерации АТФ, или в качестве субстрата для ферментов, которые синтезируют глюкозу и восстанавливают запасы гликогена.
 В период нагрузок молочная кислота диффундирует из мышечных волокон в кровоток. Этот процесс продолжается после того, как напряжение закончилось, потому что внутриклеточные концентрации молочной кислоты все еще относительно высоки. Печень поглощает молочную кислоту и преобразует ее в пировиноградную кислоту. Приблизительно 30% этих молекул пировиноградной кислоты расщепляются, обеспечивая АТФ, необходимый для превращения других молекул пировиноградной кислоты в глюкозу. Молекулы глюкозы затем выпускаются в обращение, где они поглощаются скелетными мышечными волокнами и используются для восстановления их запасов гликогена. Эта перетасовка молочной кислоты в печени и глюкозы к мышечным клеткам называется циклом Кори.
В период нагрузок молочная кислота диффундирует из мышечных волокон в кровоток. Этот процесс продолжается после того, как напряжение закончилось, потому что внутриклеточные концентрации молочной кислоты все еще относительно высоки. Печень поглощает молочную кислоту и преобразует ее в пировиноградную кислоту. Приблизительно 30% этих молекул пировиноградной кислоты расщепляются, обеспечивая АТФ, необходимый для превращения других молекул пировиноградной кислоты в глюкозу. Молекулы глюкозы затем выпускаются в обращение, где они поглощаются скелетными мышечными волокнами и используются для восстановления их запасов гликогена. Эта перетасовка молочной кислоты в печени и глюкозы к мышечным клеткам называется циклом Кори.
В период восстановления кислород легко доступен и потребность тела в кислороде остается повышенной, выше нормального уровня покоя. Восстановительный период подпитывается от АТФ. Чем больше АТФ требуется, тем больше кислорода будет необходимо. Кислородный долг или избыточное послетренировочное потребление кислорода, созданное во время физических упражнений, является тем самым количеством кислорода, которое необходимо для нормального восстановления. Скелетные мышечные волокна, которые должны восстановить АТФ, креатинфосфат и гликоген, в концентрации своих прежних уровнях и клетки печени, которые  генерируют АТФ, необходимый для превращения избыточной молочной кислоты в глюкозу, несут ответственность за большую часть дополнительного потребления кислорода. В то время, как кислородный долг восполняется, частота и глубина дыхания увеличиваются. В результате, вы будете продолжать дышать тяжело достаточно долго после прекращения интенсивных тренировок.
генерируют АТФ, необходимый для превращения избыточной молочной кислоты в глюкозу, несут ответственность за большую часть дополнительного потребления кислорода. В то время, как кислородный долг восполняется, частота и глубина дыхания увеличиваются. В результате, вы будете продолжать дышать тяжело достаточно долго после прекращения интенсивных тренировок.
Тепловые потери мышечной активности генерирует значительные объемы тепла. Когда происходит катаболическая реакция, например, при пробое гликогена или реакций гликолиза, мышечные волокна захватывают только часть выделенной энергии. Остальная высвобождается в виде тепла. Отдыхающие мышечные волокна, опирающиеся на аэробный метаболизм, захватывают около 42 % энергии, выделяемой в катаболизме. Другие 58 % согревают саркоплазму тканевой жидкости и циркулирующую кровь. Активные скелетные мышцы выпускают около 85 % тепла, необходимого для поддержания нормальной температуры тела.
Когда мышцы становятся активными, их энерго-потребление резко возрастает. Поскольку производство анаэробной энергии становится основным методом АТФ, мышечные волокна уже менее эффективно поглощают энергию. В пиковых уровнях нагрузки, только около 30 % от выделенной энергии сохраняется в виде АТФ, а остальные 70 % согревают мышцы и окружающие их ткани.
Гормоны и мышечный метаболизм. Метаболическая активность в скелетных мышечных волокнах регулируется гормонами эндокринной системы. Гормон роста из гипофиза и тестостерон (основной половой гормон у мужчин) стимулируют синтез сократительных белков и расширение скелетных мышц. Гормоны щитовидной железы поднимают скорость потребления энергии при отдыхе. Во время интенсивной физической активности, гормоны надпочечники, особенно адреналин, стимулируют мышечный обмен веществ и увеличивают продолжительность стимуляции и силу сокращения.
 Опубликовано в рубрике
Опубликовано в рубрике
классная стаья, всё интересно описано, спасибо
🙂